| К оглавлению журнала | |
|
УДК [563.14+5б1.22]:56.074 .6(470.1+470.3) |
©Коллектив авторов, 1995 |
УСЛОВИЯ ОБРАЗОВАНИЯ ПОТЕНЦИАЛЬНО НЕФТЕМАТЕРИНСКИХ ПОРОД
М.С
.Афанасьева, М.Б.Бурзин, М.В.Михайлова, Ю.Т.Кузьменко(Апрелевское отделение ВНИГНИ)Происхождение источника органического вещества - принципиальный вопрос в нефтяной геологии. Микропалеонтологический метод не только позволяет создать детальную биостратиграфию нефтегазоносных толщ, но и непосредственно изучить организмы, послужившие источником органического вещества нефтематеринских пород, а также восстановить специфические условия в бассейнах осадконакопления, способствовавшие концентрации органики.
Ревизионные исследования керна Апрелевского кернохранилища позволили поставить научно-исследовательские работы по изучению и реконструкции условий образования потенциально нефтематеринских пород франского доманика и доманикоидных отложений венда. Выбор таких контрастных объектов, как девонский доманик и вендские доманикоиды, обусловлен тем, что доманиковые отложения в Тимано-Печорском бассейне - классические палеозойские нефтематеринские породы, а верхний венд центральных районов Русской плиты - перспективный нефтегазоносный докембрийский бассейн.
В процессе развития Земли менялись бассейны осадконакопления, происходила эволюция живых организмов и их экосистем, однако анализ условий образования потенциально нефтематеринских пород франского доманика и доманикоидных отложений венда, различающихся как по возрасту и типу седиментации, так и по систематическому составу доминирующих групп организмов, может позволить выявить общие закономерности формирования этих отложений.
УСЛОВИЯ ОБРАЗОВАНИЯ ВЕНДСКИХ ДОМАНИКОИДНЫХ ПОРОД
В вендском осадочном бассейне Русской плиты слои с повышенной концентрацией органического вещества встречаются в отложениях верхнего венда (валдайский комплекс), слагающих нижнюю часть осадочного чехла, где они приурочены к нижней части редкинской серии (так называемый вендский "доманик") (Кирсанов В.В., 1968). а также к отдельным уровням поваровской серии. Вендский "доманик" - основной источник древних нефтей и битумоидов в центральных районах [2].
В результате микропалеонтологических исследований редкинских доманикоидных отложений (скв. 2, 4 Гаврилов Ям, Редкино, 1 Солигалич. Р-9 Очакова) удалось установить, что обогащенные органическим веществом прослои содержат большое число подвергшихся сильному разложению фрагментов нитчатых водорослей Eoholynia mosquensis G n i L к которым в верхней части этих отложений добавляются нитчатые водоросли Striatella coriacea A s, на фоне немногочисленных акритарх и чехлов нитчатых цианобактерий. Многие остатки инкрустированы пиритом. Однако основная масса захороненного органического вещества приурочена к пленкам сапропелеподобного вещества и микрофоссилиям Spumosina rubiginosa ( А n d r. ) - дисковидным телам, сложенным веществом, аналогичным пленкам. При сканирующей электронной микроскопии пленок и спумозин выявлено, что они "нафаршированы" фрамбоидами пирита, которые в современных и ископаемых цианобактериальных матах интерпретируются как диагенетические новообразования по остаткам колониальных серных бактерий (Krumbein W.E.. 1978; [3]).
Такая сохранность означает, что в экосистеме были условия как для продукции значительных объемов органики, так и для почти полной их деструкции. При этом практически не найдены остатки бентосных нитчатых серных бактерий, которые на западе Русской плиты развивались
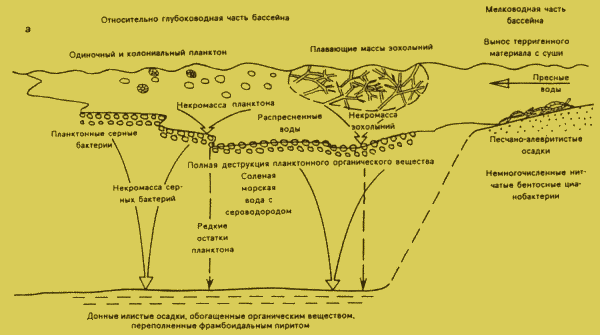
в изобилии на накапливавшемся на дне детрите (Бурзин М.Б„ 1995).Можно предложить следующую реконструкцию условий (
рис. 1, а). Располагавшийся в гумидных условиях средних или высоких широт бассейн центральных районов представлял собой гигантский эстуарий, в который со щитов сносился водами большой объем терригенного материала [I]. Следовательно, можно предположить, что его воды были стратифицированы: в придонной части располагался слой соленых морских вод, а поверхностный слой был распресненным. Такая стратификация препятствовала перемешиванию вод и поступлению кислорода в придонные слои, что должно было вызвать их сероводородное заражение. Обильная продукция была связана с цветением планктона и приносом течениями и волнами фрагментов бентосных водорослей с мелководий. Возможно, ветвящиеся водоросли Eoholynia могли образовывать большие переплетенные массы, которые подобно современной Cladophora отрывались от субстрата и существовали в подвешенном состоянии. Отмершие остатки планктона и перенесенных бентосных водорослей должны были оседать на разделе вод разной плотности, где ими питались планктонные серные бактерии, подобно тому. как это происходит в современном Черном море. В таком случае на дно в основном поступали лишь остатки собственна серных бактерий и продукты их жизнедеятельности в виде обогащенных липидами органических соединений [3]. По остаткам колоний серных бактерий, содержавших внутриклеточные гранулы серы. в процессе диагенеза формировались фрамбоиды пирита. Судя по палеогеографическим картам [1] области формирования вендского "доманика" располагались не в центральной части бассейна, а по его южной окраине, что соответствует представлениям о плотностной стратификации вод, вызванной притоком пресных вод с суши - главном условии образования обогащенных органическим веществом этих доманикоидных пород.Обнаруженные на отдельных уровнях поваровской серии (скв.
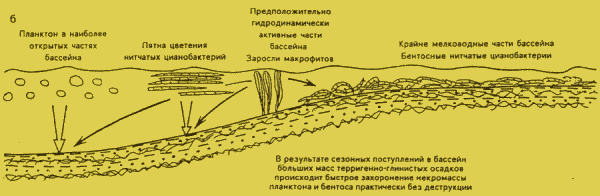
1 Гаврилов Ям) прослои, обогащенные органическим веществом, демонстрируют принципиально иную картину. Они содержат прекрасной сохранности органикостенные микрофоссилии. представляющие собой как остатки планктона (разнообразные акритархи и колонии нитчатых цианобактерий), так и бентоса (фрагменты дерновин нитчатых цианобактерий, захороненных in situ, а также ик перенесенные фрагменты и части макроскопических водорослей Vendotaenia antiqua G n i l. ). Встречаются и сапропелевые пленки с фрамбоидами пирита. Остатки, инкрустированные пиритом, не известны.По соотношению остатков планктона и бентоса, а также тафономическим данным можно восстановить фациальную гамму населения бассейна:
на крайнем мелководье обитали цианобактериальные маты, в среднеглубинных частях бассейна цианобактерий вели преимущественно планктонный образ жизни, в открытых и более глубоких водах развивался эвкариотный планктон, а в зонах с активной динамикой вод произрастали заросли вендотений (Бурзин М.Б.,
1993).Прекрасная сохранность свидетельствует, что было достаточно времени для продукции органики, но не было времени для ее деструкции или деятельность деструкторов была резко подавлена. Породы поваровской серии с повышенным количеством органических остатков имеют тонкую слоистость (даже ленточное строение), обусловленную градационной сортировкой материала, причем остатки содержатся в верхней глинистой части ритма (Бурзин М.Б.,
1989). Такое литологическое строение в сочетании с прекрасной сохранностью остатков позволяет дать следующую реконструкцию условий (см. рис, 1,б). В мелководном и судя по литологическим и геохимическим данным (Пиррус Э.Л., 1986) в ненормально-морском бассейне существовали бентосные и планктонные сообщества, цветение которых происходило в сухие сезоны. Во время влажных сезонов в бассейн поступал с суши терригенный материал, шторма могли вызывать быстрое захоронение органических остатков, делая их недоступными для деятельности деструкторов. Как удалось показать на экспериментах с современными цианобактериями (Бурзин М.Б, Орлеанский В.К, 1995), засыпание осадком или заливание раствором глины - достаточное условие для прекращения работы деструкторов, которые в культуре полностью разрушают цианобактерии за 1 мес.УСЛОВИЯ ОБРАЗОВАНИЯ ФРАНСКИХ ДОМАНИКОВЫХ ОТЛОЖЕНИЙ
Ухтинский район Тимано-Печорской нефтегазоносной провинции европейского севера России служит стратотипом доманика. Здесь доманик был впервые установлен и охарактеризован А.А.Кайзерлингом в середине
XIX в. В середине нашего столетия проблемы, связанные с домаником, описывались С.В.Тихомировым (1948, 1967), Т.И.Кушнаревой (1963), С.В.Максимовой (1970) и В.Вл. Меннером (1989). Доманиковые отложения являются нефтематеринскими породами и служат индикаторами особых палеогеографических условий бассейна седиментации. Отличительные признаки доманиковых отложений: высокое содержание органического вещества: характерные литологические фации; специфические комплексы планктонных организмов; отсутствие нормально-морского бентоса. Кроме того, существует предположение [5], что радиолярии были одним из основных источников материнского вещества нефти за счет пересыщения живых клеток липидами и возможного симбиоза с водорослями.Изученные по скважинам и в обнажениях доманиковые отложения Ухтинской части Тимано-Печорского бассейна (Афанасьева М.С.. Михайлова М.В.,
1994, 1995; [4]) представлены толщей тончайшего чередования микрозернистых, битуминозных. прослоями сильно пиритизированных кремнекарбонатных пород и кремнистых аргиллитов, а также смешанными 2-4-компонентными породами этого же состава. Набор типов пород и их количественные соотношения значительна меняются по разрезу. Соответственно изменяются соотношения различных органических остатков: тентакулитов, остракод, пелеципод, гониатитов, губок, радиолярий и акритарх. Радиолярии из собственно доманиковых отложений региона до настоящего времени практически не изучались, хотя их обильное присутствие в шлифах отмечала С.В. Максимова в 1970 г.В результате наших исследований из полного разреза доманиковых отложений впервые выделены радиолярии прекрасной сохранности, относящиеся более чем к
100 видам (рис. 2), Среди изученных радиолярий отчетливо выделяются две морфологические группы: сферические планктонные Spumellaria и иглистые бентосные Albaillellaria, отличающиеся избирательной приуроченностью к различным зонам моря и типам осадков. Морфологические особенности раковин радиолярий и характер их палеобиоценозав были теснейшим образом связаны со средой и условиями обитания и изменялись не только в пространстве при переходе от относительно глубоководных к более мелководным осадкам, но и зависели от изменения температуры вод палеоморя: преобладание в осадке иглистых форм бентосных Albaillellana на фоне мелких угнетенных сферических раковин планктонных Spumellaria может свидетельствовать об относительно холодных придонных водах палеоморей, что могло быть связано с эвстатическими колебаниями уровня доманикового палеоморя (Афанасьева М.С., Михайлова М.В.. 1994. 1995; [4]).Кремнеизвестняковые породы нижней более карбонатной части разреза отличаются сингенетичным рассеянным окремнением, высокой степенью раннедиагенетических преобразований и переслаиванием с тентакулитовыми известняками. В нижней части разреза также нередко встречаются следы ризоидов. Возможно, условия древнего моря в начале доманикоаого времени периодически способствовали скоплению растворенной кремнекислоты и жизненно важных элементов продуктов вулканической активности только в придонных относительно более прохладных водах. Это определило, с одной стороны, широкое распространение в осадках тентакупитов. остракод и гониатитов, незначительное содержание радиолярий
(50 экз. на 1 г породы), низкое видовое разнообразие и малые размеры сферических раковин планктонных Spumellana. С другой стороны, эти условия были оптимальными дли развития относительно холоднолюбивых иглистых форм бентосных Albaillellaria (до 61 %). Они. вероятно, вели придонный образ жизни, приобретая доминантное значение а ассоциации радиолярий.Средняя и верхняя части разреза отличаются повышением вверх по разрезу общей кремнистости пород, что. вероятно, было обусловлено увеличением содержания растворенной кремнекислоты в водах палеобассейна, поступавшей за счет усиления подводной вулканической активности. Произошедшие в конце доманикового времени изменения условий среды обитания сопровождались широким распространением акритарх и резким увеличением общего числа радиолярий (более
1000 экз. на 1 г породы). В это время преобладали разнообразные сферические Spumellaria (до 93 %). Численность и видовое разнообразие иглистых Aibailiellaria почти не менялись.Таким образом, анализ палеобиоценозов доманика
, включающих остракоды, гониатиты, тентакулиты, губки, радиолярии и акритархи, а также литофациальные особенности вмещающих их пород позволили выяснить условия накопления потенциально нефтематеринских осадков в обособленных зонах внешнего шельфа с глубины менее 100 м.Радиолярии, как и другие планктонные организмы, такие как коккопитофориды, фораминиферы и динофлагелляты, ассоциируются в современных морях с зонами апвеллинга [5]. Однако во время пиков кислородной недостаточности в приданных водах настоящего и прошлого радиолярии вытесняют из биоценозов нормально-морские бентосные и планктонные организмы (фораминиферы, остракоды, гониатиты. тентакулиты, губки) и ассоциируются только с акритархами.
По-видимому, только важные биологические отличия между этими группами организмов могут объяснить феномен подобного замещения [5]. Дело в том, что остракоды. гониатиты и тентакулиты размножаются только половым путем, а жизненный цикл развития губок и фораминифер включает чередование полового и бесполого поколений. Во время полового размножения гаметы и личинки опускаются на морское дно для последующего воспроизводства и развития, которое было бы невозможным при отсутствии или недостатке кислорода. Радиолярии же и акритархи размножаются путем бесполого деления клетки и не зависят от кислородных условий придонных вод.
Кроме того, цитоплазма клетки живых радиолярий наряду с различными органеллами содержит многочисленные запасные жировые включения размером 3-4 мкм и симбиотические водоросли (Петрушевская М.Г, 1986; Anderson O.R., 1992). Исходя из этого, А.Р.Ормистон [5] предположил. что основными поставщиками органики в нефтематеринских породах могли быть вместе с водорослями и радиолярии.
Предварительные результаты моделирования тела радиолярий показали, что объем цитоплазмы клетки превышает массу кремневого скелета в 100-700 раз у сферических форм и в 700-3000 раз у иглистых разностей. Массовые скопления радиолярий обогащали осадки бассейна сингенетичным органическим веществом, которое явилось результатом посмертных преобразований цитоплазмы их клеток и впоследствии могло стать одним из источников нефтегазовых углеводородов домзниковых пород.
ВЫВОДЫ
1. В верхнем венде центральных районов Русской плиты установлено два уровня накопления пород, богатых органическим веществом, с приблизительно схожими сообществами продуцентов. но принципиально различной сохранностью остатков организмов.
2. Приуроченность нефтепроявлений именно к нижнему уровню (вендскому "доманику"), вероятно, может быть объяснена "облагораживанием" органических веществ в результате жизнедеятельности деструкторов, которые разрушают целлюлозу и другие нерастворимые соединения и обогащают материал липидами.
3. Для образования нефтематеринских пород важны не только благоприятные условия среды, обеспечивающие продукцию большого количества органики, но и последующая бактериальная деструкция, для чего необходима определенная временная задержка до захоронения в осадке.
4. Ухтинская часть бассейна доманикового осадконакопления представляла собой обособленную экологическую зону внешнего шельфа, где могли происходить существенные изменения глубины в связи с эвстатическими колебаниями уровня моря. а максимальные отметки до 100 м, вероятнее всего, существовали лишь в отдельных частях палеобассейна и в короткие интервалы времени.
5. Преобладание в осадке иглистых форм бентосных Albaillellaria на фоне мелких угнетенных сферических раковин планктонных Spumellaria может свидетельствовать об относительно холодных приданных водах палеоморей, что могло быть связано с эвстатическими колебаниями уровня доманикового палеоморя.
6, Накопление осадков в условиях бескислородной среды, способствующей сохранению органического вещества, оптимально для формирования нефтематеринских пород, а предположение, что радиолярии в значительной степени способствуют увеличению органики в осадках, достаточно вероятно.
ЛИТЕРАТУРА
1.
Аксенов Е.М. Венд Восточно-Европейской платформы //Вендская система. - М„ 1985. -Т. 2. - С.3-34.2.
Баженова O.K., Арефьев ОД., Соколов Б.А. Генетические особенности нефтей верхнего протерозоя //Докл. РАН. - 1994. - Т.337. -№ 3. - С.371-375.3.
Роль бентосных микробных сообществ в формировании нефтематеринского потенциала / С.И.Жмур, О.К.Бзженова, М.Б.Бурзин. В.М.Горленко, А.Ю.Розанов. Б.А.Соколов //История нефти в осадочных бассейнах. - М.. 1994. -С.35-48.4 Afanasieva M.S., Mikhailova M.V.
Domanik Radiolarians as bioindicators of conditions of sedimentation //Seventh meeting of the International Association of Radiolarian Paleontoiogists (INTER-RAD VII). Osaka (Japan), October 20-24. 1994. // Osaka City University. Abstracts. - 1994. - P.3.5. Ormiston A.R. The association of radiolarians with hydrocarbon source rocks //Micropaleontology. Special Publication. - 1992. - № 6. - P.9-16.
Рис.1. РЕКОНСТРУКЦИЯ ПАЛЕОЭКОЛОГИЧЕСКИХ УСЛОВИЙ В БАССЕЙНЕ ЦЕНТРАЛЬНЫХ РАЙОНОВ РУССКОЙ ПЛИТЫ ВО ВРЕМЯ НАКОПЛЕНИЯ ОСАДКОВ В НАЧАЛЕ РЕДКИНСКОГО (а) И В ПОВАРОВСКОЕ ВРЕМЯ (б) ПОЗДНЕГО ВЕНДА
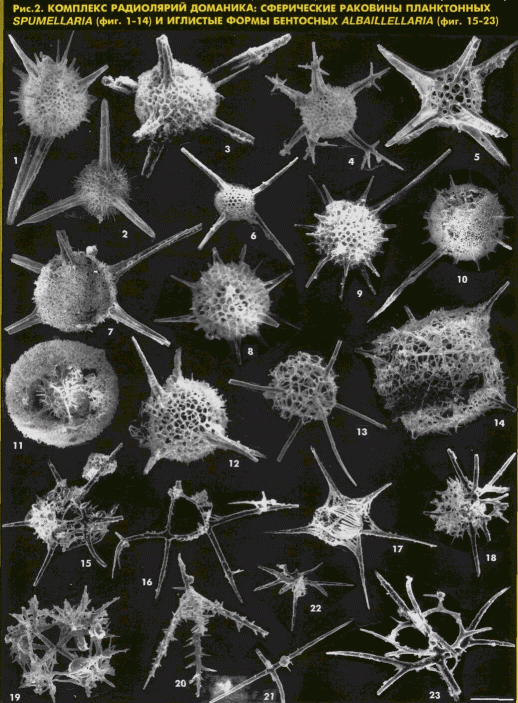
Фиг.
1, 2, 5-12, 14, 15, 17-22: скв. 1003 Шуда-Яг; фиг. 3, 4, 13, 16, 23: р. Лыайёль, обнажение 19, образец 1906.1. Entactinosphaera assidera N a z
а г о v; образец 73 (интервал 71,4-71,9 м); АО-94/8821;СЭМ, штрих - 72 мкм.2- Entactinosphaera grandis N
а z а г о v;образец 73 (интервал 71,4-71,9 м): АО-94/8817;СЭМ, штрих - 145 мкм.3- Spongentactinia somphorhips (Foreman):
АО-94/9705; СЭМ, штрих - 59 мкм,4. Entactinia sp.33:
АО-94/9512; СЭМ, штрих - 72 мкм.5. Entactinia micula Foreman;
образец 68 (интервал 73,0-73,3 м): АО-94/4727; СЭМ, штрих - 48 мкм.6. Entactinia paula Foreman;
образец 78 (интервал 68,9-69,3 м): АО-94/4919; СЭМ, штрих - 108 мкм.7. Spongentactinella sp.56;
образец 72 (интервал 71,9-72,4 м): АО-94/8514; СЭМ, штрих - 54 мкм.8. Astroentactinia biaciculata N
а z а г о v;образец 72 (интервал 71,9-72,4 м); АО-94/8517;СЭМ, штрих - 59 мкм.9. Astroentactinia paronae ( H i n d e );
образец 72 (интервал 71,9-72,4 м): АО-94/8627;СЭМ, штрих - 86 мкм.10. Helioentactinia sp.60;
образец 73 (интервал 71,4-71,9 м): АО-94/8728; СЭМ, штрих - 81 мкм.11. Somphoentactinia sp.62;
образец 68 (интервал 73,0-73,3 м): АО-94/4710; СЭМ, штрих - 108 мкм.12. Haplentactinia pantosompha (Foreman);
образец 68 (интервал 73,0-73,3 м): АО-94/4725;СЭМ, штрих - 54 мкм.13. Tetragregnon sp.3:
АО-94/9429; СЭМ, штрих - 72 мкм.14. Polyentactinia sp.11;
образец 68 (интервал 73,0-73,3 м): АО-94/4722; СЭМ, штрих - 65 мкм.15. Ceratoikiscum delicatum
С h e n g ;образец 29 (интервал 105-106 м): АО-94/4503;СЭМ, штрих - 87 мкм.16. Ceratoikiscum sp.106:
АО-94/9835; СЭМ, штрих - 87 мкм.17. Neoholoeciscum avimexpectans (Dcflandrc);
образец 68 (интервал 73,5-73,3 м): АО-94/4724;СЭМ, штрих - 100 мкм.18. Xiphocabrium(?) sp,97;
образец 68 (интервал 73,0-73,3 м): АО-94/4724; СЭМ, штрих - 100 мкм.19. Palaeothalomnus sp-86;
образец 73 (интервал 71,4-71,9 м): АО-94/8713; СЭМ, штрих - 72 мкм-20. Palaeoscenidium. cladophorum D e f 1 a n d r e ;
образец 72 (интервал 71,9-72,4 м): АО-94/5133;СЭМ, штрих - 87 мкм.21. Palacantholithus sp-89;
образец 8 (интервал 120,9-124,0 м): АО-94/4028; СЭМ, штрих - 130 мкм.22. Xiphocladiella(?) sp-93;
образец 28 (интервал 106-107 м): АО-94/4218; СЭМ, штрих - 130 мкм.23. Xiphocladiella(?) sp.92:
АО-94/9834; СЭМ, штрих - 72 мкм.